ДОКЛАДЫ АКАДЕМИИ НАУК, 2007, том 417, № 4, с. 550-552
БИОХИМИЯ, БИОФИЗИКА, МОЛЕКУЛЯРНАЯ БИОЛОГИЯ
УДК 577.21:577.171.7:577.175.823
ИЗМЕНЕНИЕ ЭКСПРЕССИИ ГЕНОВ ВНУТРИКЛЕТОЧНЫХ СИГНАЛЬНЫХ ПУТЕЙ В ГИППОКАМПЕ КРЫС ПОД ДЕЙСТВИЕМ ПЕПТИДА СЕМАКС
© 2007 г. Т. Ю. Агапова, Я. В. Агниуллин, Д. Н. Силачев, М. И. Шадрина, П. А. Сломинский, С. И. Шрам, С. А. Лимборская, академик Н. Ф. Мясоедов
Поступило 06.07.2007 г.
АКТГ [4-7] - самый короткий ^концевой фрагмент адренокортикотропного гормона, способный влиять на поведение животных [1]. Для создания ноотропных лекарств на основе фрагмента АКТГ [4-7] был разработан и испытан ряд его синтетических аналогов [2]. В лабораториях ИМГ РАН и МГУ создали пептид Met-Glu-His-Phe-Pro-Gly-Pro, названный семаксом. Показано, что семакс способствует выживанию клеток мозга при гипоксии, усиливает избирательное внимание и консолидацию памятного следа [3]. Лекарственные препараты на его основе применяются для лечения черепно-мозговых травм, инсульта, заболеваний зрительного нерва, в постреанимационном периоде [4, 5].
Механизм действия семакса во многом остается неясным [6, 7]. Ранее было показано увеличение экспрессии генов нейротрофинов и их специфических рецепторов под действием семакса [2, 8-10]. Для изучения механизма действия семакса на уровне транскриптома нами был проведен расши-
ренный поиск генов, изменяющих экспрессию иод влиянием этого пептида.
В опыте использовали самцов крыс линии Ви-стар массой 190-210 г. Двум группам животных (по 5 особей) в течение 10 дней дважды в день ин-траназально вводили воду. На одиннадцатые сутки "контрольной" группе закапали воду, а "опытной" - водный раствор семакса (50 мкг/кг массы). Через час животных декапитировали и из гиппо-кампа выделяли тотальную РНК, на основе которой получали одноцепочечную кДНК. Подробнее выбор экспериментальных групп животных и методы получения кДНК описаны нами ранее [10].
Влияние семакса на экспрессию 84 генов, представляющих 18 различных путей передачи сигнала, было проанализировано с помощью панели генов "Signal Transduction PathwayFinder" ("RT2 Profiler™ PCR Array", "SuperArray", США) с использованием ПЦР в реальном времени. Реакции повторяли трижды для каждого образца ткани. Полученные значения пороговых циклов реакций
Wnt-путь
Изменение уровня экспрессии, число раз 1.50г 1.25-
123456789 10 11 Ген
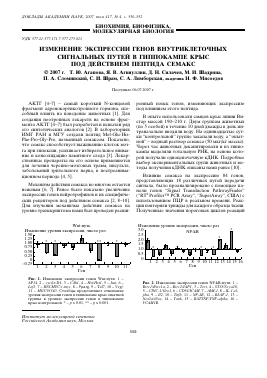
Рис. 1. Изменение экспрессии генов Wnt-пути. 1 -АР14, 2 - сусПп D1, 3 - Cdh1, 4 - Ш^Ш^, 5 - Jun, 6 -Lef1, 7 - RNCMYC/c-myc, 8 - Рраг^ 9 - ТсП, 10 - Vegf, 11 - МСС93340. Столбцы представляют отношение уровня экспрессии генов в гиппокампе крыс опытной группы к уровню экспрессии генов в гиппокампе крыс контрольной. * -р < 0.01, ** -р < 0.001.
Изменение уровня экспрессии, число раз 3 0г *
3'5Г NF-kB
2.0 1.5 1.0
0.5
а
и У У ^ У 1 и * и
Jk
_|_I_I_I_I_I_I_I_I_I_I_I_I_I_I_1
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 Ген
Рис. 2. Изменение экспрессии генов NF-kB-пути. 1 -Bircl/Bircla, 2 - Birc2/IAP1, 3 - Tert, 4 - ST38/Scya20, 5 - CINC-1/Gro1, 6 - CD54/ICAM, 7 - AIM-1, 8 - IL-1 alpha, 9 - Il2, 10 - Tnfb, 11 - NF-kB, 12 - RL/IF-1, 13 -Nos2a/iNos, 14 - Tank, 15 - RATTNF/TNF-alpha, 16 -VCAM1B.
0
Институт молекулярной генетики Российской Академии наук, Москва
ИЗМЕНЕНИЕ ЭКСПРЕССИИ ГЕНОВ
551
Таблица 1. Падение экспрессии генов через час после введения препарата семакс
Таблица 2. Возрастание экспрессии генов через один час после введения препарата семакс
Название гена Название пути передачи сигнала Изменение экспрессии
Nab2: Ngfi-A связывающий белок 2 Митогенный 0.44*
Ccndl: Циклин D1 Wnt 0.45*
Cdhl: Кадгерин 1 То же 0.31*
Pparg: гамма субъединица рецептора, активируемого пролифера-тором пероксисом » 0.01**
Vegfa: васкулярный эн-дотелиальный фактор роста A » 0.22*
Ptchl: гомолог гена Patched 1 дрозофилы Hedgehog 0.48*
Arf/INK4A: ингибитор циклин-зависимой киназы 2A TGF-p 0.42*
Mmp7: металлопротеи-наза матрикса 7 Путь фосфоинози-тол-3 киназы/AKT 0.21*
Birc3: ингибитор белка апоптоза 1 NF-кВ 0.28*
Lta: лимфотоксин A То же 0.38*
Nfkbia: альфа субъединица ингибитора NF-кВ » 0.49*
Tnfsf6: надсемейство факторов некроза опухоли (лиганд) 6 NFAT 0.12*
Mmp10: металлопроте-иназа матрикса 10 Jak-Stat 0.20**
Cdk2: циклин-зависи-мая киназа 2 Андрогеновый 0.37*
Tmepai: трансмембранный индуцируемый ан-дрогенами белок простаты Андрогеновый 0.16*
Rbpl: клеточный рети-нол-связывающийся белок 1 Ретиноловой кислоты 0.20**
Tfrc: рецептор транс-феррина Кальция и проте-инкиназы C 0.20*
Lep: лептин Инсулиновый 0.35*
Selp: тромбоцитарный селектин ЛПНП 0.15*
Название гена Путь передачи Изменение
сигнала экспрессии
ЮС291936: белок, гомо- Hedgehog 2.64*
логичный к hedgehog-взаи-
модействующему белку
Hspb1: 27 кД белок тепло- Стрессовый 4.94*
вого шока 1
Nos2: индуцибельная N0 NF-кВ 2.03**
синтаза 2
Ги/ член надсемейства фак- NF-кВ 2.90*
торов некроза опухоли 2
Cd5: CD5 антиген NFAT 2.31*
Схс19: хемокиновый (С-Х-С Jak-Stat 4.63*
мотив) лиганд 9
Вгса1: ген рака молочной Эстрогеновый 2.73*
железы 1
Сс12: хемокиновый (С-С ЛПНП 2.03*
мотив) лиганд 2
Примечание. Здесь и в табл. 2. Показано, во сколько раз изменился уровень экспрессии данного гена в ткани гиппокампа крыс "опытной" группы по отношению к уровню экспрессии того же гена в ткани гиппокампа крыс "контрольной" группы. * p < 0.01, ** p < 0.001.
(Ct) были нормализованы и проанализированы с помощью программного обеспечения "Relative Expression Software Tool - 384", версия 2 [11].
Полученные результаты приведены в табл. 1 и 2. Более, чем в 2 раза, изменили экспрессию 27 из 84 исследованных генов (32.1%). При этом у 19 генов (22.6%) уровень экспрессии в гиппокампе крыс, которым вводили пептид, снизился. У 8 генов (9.5%) экспрессия возросла. Наиболее достоверные изменения уровня экспрессии наблюдались у генов Pparg (экспрессия уменьшилась более, чем в сто раз), Tmepai (экспрессия уменьшилась в 6.5 раз), Apt1Lg1/Fasl (падение экспрессии в 8.1 раза), Rbp1 (падение экспрессии в 5 раз). Сильно изменился уровень экспрессии Vegf (упал в 4.5 раза), Mig/Scyb9 (возрос в 4.6 раз),Mmp10 (упал в 5.1 раза), Trfr (упал в 5 раз) и еще ряда генов.
Наиболее информативными для анализа представляются данные, отражающие действие пептида на Wnt-путь (значительно уменьшилась экспрессия 36.4% генов (см. рис. 1)), и NF-кВ-путь (уровень экспрессии 31.3% генов изменился в различных направлениях (см. рис. 2)). Мы видим, что действие семакса на разные звенья внутриклеточных сигнальных путей дифференцированно. Это указывает на возможное участие этого пептида в регуляции процессов, контролируемых исследованными внутриклеточными путями передачи сигнала. Как известно, белки Wnt-пути передачи сигнала участвуют в процессах дифференциров-ки, деления, ориентации, адгезии, выживания и апоптоза в клетках. В частности, они способны угнетать апоптоз и увеличивать способность нейронов или нейрональных культур клеток к выживанию путем активации NF-кВ-пути [12].
ДОКЛАДЫ АКАДЕМИИ НАУК том 417 < 4 2007
552
АГАПОВА и др.
Под влиянием семакса в ткани гиппокампа значительно изменили свою экспрессию гены большинства путей передачи сигнала, представленных в анализируемой панели генов. Необходимы дальнейшие исследования для подтверждения полученных данных и установления возможного механизма действия пептида семакса.
Данное исследование проводилось при поддержке грантов Российского фонда фундаментальных исследований (грант 06-04-49368-a, грант 04-04-08117-офи(а), Программ РАН "Молекулярная и клеточная биология", "Фундаментальные науки - медицине", государственных контрактов с Министерством образования и науки РФ.
СПИСОК ЛИТЕРАТУРЫ
1. De Wied D, Witter A., Greven HM. // Biochem. Pharmacol. 1975. V. 24. № 16. P. 1463-1468.
2. Shadrina M.I., Dolotov O.V., Grivennikov I.A. et al. // Neurosci. Lett. 2001. V. 308. № 2. P. 115-118.
3. Ашмарин И.П., Незавибатько ВН., Мясоедов Н.Ф. и др. // ЖВНД. 1997. Т. 47. № 2. С. 420-430.
4. Kypышева НИ, Шпак А.А., Иойлева Е.Э. и др. // Вестн. офтальмологии. 2001. Т. 117. № 4. С. 5-8.
5. Мясоедов Н.Ф, Скворцова ВИ, Насонов ЕЛ. и др. // Журн. неврологии и психиатрии им. С.С. Корсакова. 1999. Т. 99. № 5. С. 15-19.
6. Шевченко КВ., Нагаев И.Ю., Алфеева Л.Ю. и др. // Биоорган. химия. 2006. Т. 32. № 1. С. 64-70.
7. Eremin K.O, Kudrin V.S., Saransaari P. et al. // Neuro-chem. Res. 2005. V. 30. № 12. P. 1493-1500.
8. Dolotov O.V., Karpenko E.A., Inozemtseva L.S. et al. // Brain Res. 2006. V. 1117. № 1. С. 54-60.
9. Zolotarev YA, Dolotov O.V., Inozemtseva L.S. et al. // Amino Acids. 2006. V. 30. № 4. P. 403-408.
10. Agapova T.Y., Agniullin Y.V., Shadrina M.I. et al. Neurosci. Lett. 2007. V. 417. № 2. P. 201-205.
11. Pfaffl M.W, Tichopad A., Prgomet C, Neuvians TP. // Biotechnol. Lett. 2004. V. 26. № 6. P. 509-515.
12. Li F, Chong ZZ, Maiese K. // Curr. Neurovasc. Res. 2005. V. 2. № 4. P. 331-340.
ДОКЛАДЫ АКАДЕМИИ НАУК том 417 < 4 2007
Для дальнейшего прочтения статьи необходимо приобрести полный текст. Статьи высылаются в формате PDF на указанную при оплате почту. Время доставки составляет менее 10 минут. Стоимость одной статьи — 150 рублей.